The Evolution of Populations Reading Guide Answers
Talk over the ways populations evolve
All life on Earth is related. Evolutionary theory states that humans, beetles, plants, and bacteria all share a common antecedent, simply that millions of years of development have shaped each of these organisms into the forms seen today. Scientists consider evolution a primal concept to understanding life. Natural selection is one of the near dominant evolutionary forces. Natural choice acts to promote traits and behaviors that increase an organism's chances of survival and reproduction, while eliminating those traits and behaviors that are to the organism'due south detriment. But natural selection tin simply, every bit its name implies, select—it cannot create. The introduction of novel traits and behaviors falls on the shoulders of another evolutionary forcefulness—mutation. Mutation and other sources of variation among individuals, also as the evolutionary forces that human action upon them, alter populations and species. This combination of processes has led to the world of life we see today.
Learning Objectives
- Describe how population genetics is used in the study of the evolution of populations
- Define the Hardy-Weinberg principle and discuss its importance
- Describe the different types of variation in a population
- Explain the different ways natural selection tin can shape populations
Population Genetics
Recall that a gene for a particular character may have several alleles, or variants, that code for different traits associated with that character. For instance, in the ABO blood type system in humans, three alleles determine the detail blood-type protein on the surface of red blood cells. Each individual in a population of diploid organisms can only carry two alleles for a particular gene, but more two may be present in the individuals that make up the population. Mendel followed alleles as they were inherited from parent to offspring. In the early twentieth century, biologists in a discipline known as population genetics began to study how selective forces change a population through changes in allele and genotypic frequencies.
The allele frequency (or gene frequency) is the rate at which a specific allele appears within a population. Until now we have discussed evolution as a alter in the characteristics of a population of organisms, but backside that phenotypic change is genetic change. In population genetics, the term evolution is defined as a change in the frequency of an allele in a population. Using the ABO blood type system as an example, the frequency of one of the alleles, I A, is the number of copies of that allele divided past all the copies of the ABO cistron in the population. For example, a study in Jordan[i] plant a frequency of I A to exist 26.one percent. The I Band I 0 alleles made up 13.4 percent and threescore.5 percent of the alleles respectively, and all of the frequencies added up to 100 percent. A alter in this frequency over time would constitute evolution in the population.
The allele frequency within a given population can change depending on environmental factors; therefore, certain alleles become more widespread than others during the procedure of natural selection. Natural choice can modify the population's genetic makeup; for case, if a given allele confers a phenotype that allows an private to amend survive or have more offspring. Because many of those offspring will too carry the beneficial allele, and often the corresponding phenotype, they will accept more offspring of their own that also carry the allele, thus, perpetuating the cycle. Over time, the allele will spread throughout the population. Some alleles volition quickly go fixed in this way, meaning that every individual of the population will conduct the allele, while detrimental mutations may be swiftly eliminated if derived from a ascendant allele from the gene pool. The factor puddle is the sum of all the alleles in a population.
Sometimes, allele frequencies inside a population alter randomly with no advantage to the population over existing allele frequencies. This phenomenon is called genetic drift. Natural selection and genetic drift usually occur simultaneously in populations and are not isolated events. It is hard to determine which process dominates because it is often nearly impossible to make up one's mind the cause of change in allele frequencies at each occurrence. An result that initiates an allele frequency modify in an isolated office of the population, which is not typical of the original population, is called the founder effect. Natural selection, random drift, and founder effects tin can lead to pregnant changes in the genome of a population.
Hardy-Weinberg Principle of Equilibrium
In the early twentieth century, English mathematician Godfrey Hardy and High german physician Wilhelm Weinberg stated the principle of equilibrium to describe the genetic makeup of a population. The theory, which subsequently became known as the Hardy-Weinberg principle of equilibrium, states that a population'south allele and genotype frequencies are inherently stable— unless some kind of evolutionary strength is interim upon the population, neither the allele nor the genotypic frequencies would change. The Hardy-Weinberg principle assumes weather with no mutations, migration, emigration, or selective pressure for or against genotype, plus an infinite population; while no population can satisfy those conditions, the principle offers a useful model against which to compare real population changes.
Working under this theory, population geneticists represent unlike alleles as different variables in their mathematical models. The variable p, for example, often represents the frequency of a particular allele, say Y for the trait of yellow in Mendel's peas, while the variable q represents the frequency of y alleles that confer the color light-green. If these are the but two possible alleles for a given locus in the population, p + q = 1. In other words, all the p alleles and all the q alleles make up all of the alleles for that locus that are institute in the population.
Simply what ultimately interests virtually biologists is not the frequencies of dissimilar alleles, but the frequencies of the resulting genotypes, known as the population'southward genetic structure, from which scientists tin can surmise the distribution of phenotypes. If the phenotype is observed, only the genotype of the homozygous recessive alleles can be known; the calculations provide an approximate of the remaining genotypes. Since each individual carries two alleles per gene, if the allele frequencies (p and q) are known, predicting the frequencies of these genotypes is a uncomplicated mathematical calculation to make up one's mind the probability of getting these genotypes if two alleles are drawn at random from the gene pool. So in the above scenario, an individual pea plant could be pp (YY), and thus produce yellow peas; pq (Yy), besides yellow; or qq (yy), and thus producing green peas (Effigy 1). In other words, the frequency of pp individuals is only p2; the frequency of pq individuals is 2pq; and the frequency of qq individuals is q2. And, once again, if p and q are the only two possible alleles for a given trait in the population, these genotypes frequencies volition sum to i: p2+ 2pq + q2 = one.
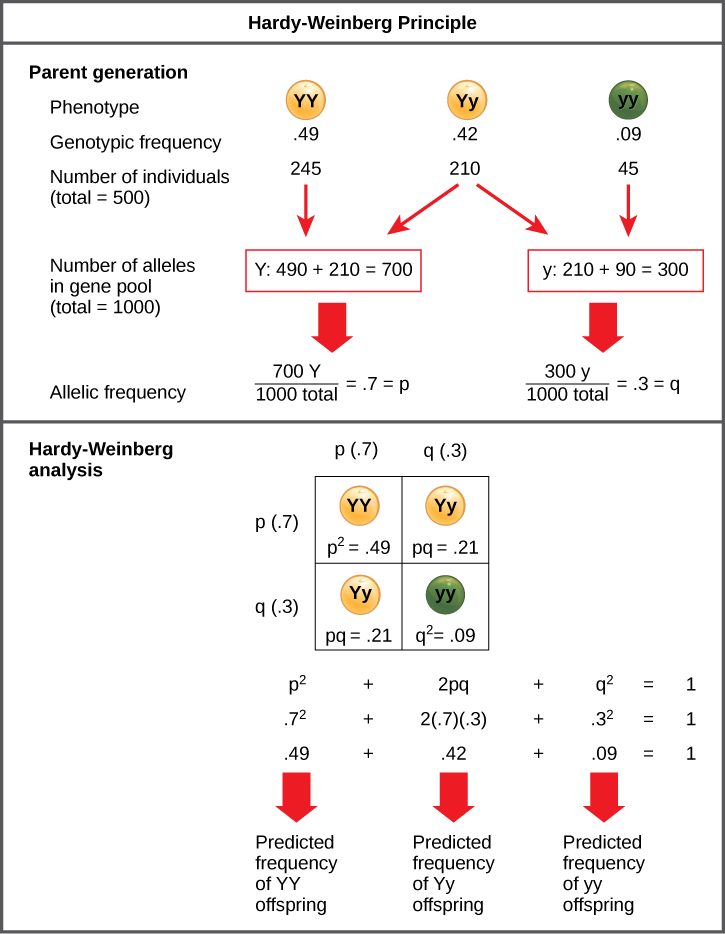
Figure 1. When populations are in the Hardy-Weinberg equilibrium, the allelic frequency is stable from generation to generation and the distribution of alleles can be adamant from the Hardy-Weinberg equation. If the allelic frequency measured in the field differs from the predicted value, scientists tin can brand inferences virtually what evolutionary forces are at play.
Practice Question
In plants, violet flower colour (V) is dominant over white (five). If p = 0.8 and q = 0.2 in a population of 500 plants, how many individuals would you look to exist homozygous dominant (VV), heterozygous (Vv), and homozygous recessive (vv)? How many plants would y'all expect to have violet flowers, and how many would take white flowers?
Show Answer
The expected distribution is 320 VV, 160Vv, and 20 vv plants. Plants with VV or Vv genotypes would have violet flowers, and plants with the vv genotype would have white flowers, so a total of 480 plants would be expected to have violet flowers, and xx plants would have white flowers.
In theory, if a population is at equilibrium—that is, there are no evolutionary forces acting upon it—generation after generation would accept the same cistron puddle and genetic structure, and these equations would all agree truthful all of the time. Of course, even Hardy and Weinberg recognized that no natural population is immune to evolution. Populations in nature are constantly changing in genetic makeup due to migrate, mutation, possibly migration, and selection. Equally a upshot, the only fashion to determine the exact distribution of phenotypes in a population is to exit and count them. But the Hardy-Weinberg principle gives scientists a mathematical baseline of a non-evolving population to which they can compare evolving populations and thereby infer what evolutionary forces might be at play. If the frequencies of alleles or genotypes deviate from the value expected from the Hardy-Weinberg equation, and then the population is evolving.
Genetic Variation and Drift

Figure 2. The distribution of phenotypes in this litter of kittens illustrates population variation. (credit: Pieter Lanser)
Individuals of a population frequently display different phenotypes, or express unlike alleles of a particular cistron, referred to every bit polymorphisms. Populations with two or more variations of particular characteristics are called polymorphic. The distribution of phenotypes among individuals, known as the population variation, is influenced by a number of factors, including the population's genetic construction and the environment (Figure 2). Agreement the sources of a phenotypic variation in a population is important for determining how a population will evolve in response to unlike evolutionary pressures.
Genetic Variance
Natural selection and some of the other evolutionary forces can only act on heritable traits, namely an organism's genetic lawmaking. Considering alleles are passed from parent to offspring, those that confer beneficial traits or behaviors may be selected for, while deleterious alleles may be selected confronting. Acquired traits, for the most role, are not heritable. For example, if an athlete works out in the gym every day, building up muscle strength, the athlete's offspring will not necessarily abound up to be a trunk builder. If there is a genetic basis for the ability to run fast, on the other hand, this may be passed to a child.
Before Darwinian development became the prevailing theory of the field, French naturalist Jean-Baptiste Lamarck theorized that acquired traits could, in fact, exist inherited; while this hypothesis has largely been unsupported, scientists have recently begun to realize that Lamarck was not completely wrong. Visit this site to larn more than.
Heritability is the fraction of phenotype variation that tin be attributed to genetic differences, or genetic variance, among individuals in a population. The greater the hereditability of a population's phenotypic variation, the more than susceptible it is to the evolutionary forces that act on heritable variation.
The diversity of alleles and genotypes within a population is called genetic variance. When scientists are involved in the breeding of a species, such every bit with animals in zoos and nature preserves, they attempt to increase a population's genetic variance to preserve as much of the phenotypic diversity as they can. This also helps reduce the risks associated with inbreeding, the mating of closely related individuals, which can accept the undesirable effect of bringing together deleterious recessive mutations that can cause abnormalities and susceptibility to affliction. For example, a disease that is acquired by a rare, recessive allele might be in a population, only information technology volition only manifest itself when an private carries two copies of the allele. Because the allele is rare in a normal, healthy population with unrestricted habitat, the hazard that 2 carriers will mate is low, and even then, only 25 pct of their offspring will inherit the disease allele from both parents. While it is likely to happen at some point, it will not happen frequently plenty for natural pick to exist able to swiftly eliminate the allele from the population, and as a consequence, the allele will be maintained at low levels in the gene pool. However, if a family of carriers begins to interbreed with each other, this will dramatically increase the likelihood of two carriers mating and somewhen producing diseased offspring, a miracle known equally inbreeding depression.
Changes in allele frequencies that are identified in a population can shed low-cal on how it is evolving. In improver to natural selection, there are other evolutionary forces that could be in play: genetic migrate, gene menstruum, mutation, nonrandom mating, and environmental variances.
Genetic Drift
The theory of natural selection stems from the observation that some individuals in a population are more probable to survive longer and have more offspring than others; thus, they will pass on more of their genes to the adjacent generation. A big, powerful male gorilla, for case, is much more likely than a smaller, weaker one to become the population's silverback, the pack'southward leader who mates far more than the other males of the group. The pack leader will father more offspring, who share half of his genes, and are likely to besides grow bigger and stronger similar their father. Over time, the genes for bigger size will increase in frequency in the population, and the population will, as a event, grow larger on average. That is, this would occur if this particular option pressure, or driving selective force, were the only one acting on the population. In other examples, better camouflage or a stronger resistance to drought might pose a pick pressure.
Another way a population's allele and genotype frequencies can change is genetic migrate (Effigy 3), which is simply the result of risk. By hazard, some individuals will have more offspring than others—not due to an reward conferred by some genetically-encoded trait, simply but because ane male happened to be in the right identify at the right time (when the receptive female walked by) or because the other ane happened to be in the wrong identify at the wrong time (when a fox was hunting).

Figure 3. Click for a larger image. Genetic drift in a population can pb to the emptying of an allele from a population past chance. In this example, rabbits with the brown glaze color allele (B) are dominant over rabbits with the white coat color allele (b). In the first generation, the 2 alleles occur with equal frequency in the population, resulting in p and q values of .5. Only one-half of the individuals reproduce, resulting in a second generation with p and q values of .7 and .3, respectively. Only two individuals in the second generation reproduce, and past chance these individuals are homozygous dominant for brown coat color. As a result, in the third generation the recessive b allele is lost.
Practise Question
Do you remember genetic drift would happen more speedily on an isle or on the mainland?
Bear witness Respond
Genetic drift is likely to occur more rapidly on an island where smaller populations are expected to occur.
Small populations are more susceptible to the forces of genetic drift. Large populations, on the other hand, are buffered against the furnishings of chance. If one private of a population of 10 individuals happens to die at a young age earlier it leaves any offspring to the adjacent generation, all of its genes—1/10 of the population'due south gene puddle—will exist of a sudden lost. In a population of 100, that'southward only i percent of the overall genetic pool; therefore, it is much less impactful on the population's genetic structure.
Watch this animation of random sampling and genetic drift in activity:

Bottleneck Effect

Figure 4. A risk event or catastrophe can reduce the genetic variability inside a population.
Genetic drift can also be magnified past natural events, such as a natural disaster that kills—at random—a large portion of the population. Known every bit the bottleneck issue, it results in a large portion of the genome suddenly being wiped out (Figure four). In i fell dive, the genetic construction of the survivors becomes the genetic structure of the entire population, which may be very dissimilar from the pre-disaster population.
Founder Event
Another scenario in which populations might experience a strong influence of genetic drift is if some portion of the population leaves to start a new population in a new location or if a population gets divided by a physical barrier of some kind. In this situation, those individuals are unlikely to be representative of the entire population, which results in the founder effect. The founder outcome occurs when the genetic structure changes to match that of the new population's founding fathers and mothers. The founder effect is believed to have been a key factor in the genetic history of the Afrikaner population of Dutch settlers in South Africa, equally evidenced by mutations that are common in Afrikaners just rare in most other populations. This is likely due to the fact that a college-than-normal proportion of the founding colonists carried these mutations. As a consequence, the population expresses unusually high incidences of Huntington's disease (Hard disk) and Fanconi anemia (FA), a genetic disorder known to crusade blood marrow and congenital abnormalities—fifty-fifty cancer.
Watch this short video to learn more nigh the founder and clogging effects. Note that the video has no audio.
Testing the Bottleneck Effect
Question: How do natural disasters affect the genetic construction of a population?
Background: When much of a population is suddenly wiped out by an convulsion or hurricane, the individuals that survive the event are usually a random sampling of the original group. As a result, the genetic makeup of the population tin can modify dramatically. This miracle is known as the bottleneck effect.
Hypothesis: Repeated natural disasters will yield dissimilar population genetic structures; therefore, each time this experiment is run, the results will vary.
Test the hypothesis: Count out the original population using unlike colored beads. For example, cherry-red, blueish, and yellow chaplet might represent cerise, blue, and xanthous individuals. Afterwards recording the number of each individual in the original population, place them all in a bottle with a narrow cervix that will simply let a few beads out at a time. And then, pour 1/three of the bottle's contents into a basin. This represents the surviving individuals afterward a natural disaster kills a bulk of the population. Count the number of the different colored beads in the basin, and tape it. Then, place all of the beads dorsum in the bottle and echo the experiment four more than times.
Clarify the data: Compare the five populations that resulted from the experiment. Do the populations all contain the same number of unlike colored chaplet, or practice they vary? Remember, these populations all came from the same exact parent population.
Course a conclusion: Most likely, the five resulting populations will differ quite dramatically. This is because natural disasters are not selective—they kill and spare individuals at random. Now call back about how this might affect a real population. What happens when a hurricane hits the Mississippi Gulf Declension? How exercise the seabirds that alive on the embankment fare?
Cistron Flow

Figure v. Gene menses can occur when an individual travels from one geographic location to another.
Another important evolutionary strength is factor flow: the menstruum of alleles in and out of a population due to the migration of individuals or gametes (Figure 5). While some populations are adequately stable, others experience more flux. Many plants, for instance, transport their pollen far and broad, past wind or by bird, to pollinate other populations of the same species some distance away. Even a population that may initially appear to exist stable, such as a pride of lions, can experience its off-white share of immigration and emigration as developing males leave their mothers to seek out a new pride with genetically unrelated females. This variable flow of individuals in and out of the grouping non but changes the gene structure of the population, but information technology can also introduce new genetic variation to populations in dissimilar geological locations and habitats.
Mutation
Mutations are changes to an organism's Deoxyribonucleic acid and are an important commuter of diversity in populations. Species evolve because of the accumulation of mutations that occur over time. The advent of new mutations is the most mutual way to introduce novel genotypic and phenotypic variance. Some mutations are unfavorable or harmful and are quickly eliminated from the population past natural pick. Others are benign and will spread through the population. Whether or not a mutation is benign or harmful is adamant by whether it helps an organism survive to sexual maturity and reproduce. Some mutations practice not practise anything and can linger, unaffected by natural selection, in the genome. Some can take a dramatic result on a gene and the resulting phenotype.
Nonrandom Mating
If individuals nonrandomly mate with their peers, the result can be a changing population. There are many reasons nonrandom mating occurs. One reason is simple mate choice; for example, female peahens may prefer peacocks with bigger, brighter tails. Traits that lead to more matings for an individual become selected for past natural selection. One common course of mate option, called assortative mating, is an individual'southward preference to mate with partners who are phenotypically similar to themselves.
Another crusade of nonrandom mating is physical location. This is specially true in large populations spread over large geographic distances where not all individuals will have equal access to one another. Some might exist miles apart through forest or over rough terrain, while others might live immediately nearby.
Environmental Variance

Figure half dozen. The sex of the American alligator (Alligator mississippiensis) is adamant by the temperature at which the eggs are incubated. Eggs incubated at 30°C produce females, and eggs incubated at 33°C produce males. (credit: Steve Hillebrand, USFWS)
Genes are not the simply players involved in determining population variation. Phenotypes are also influenced by other factors, such every bit the environment (Figure 6). A beachgoer is likely to have darker skin than a urban center dweller, for case, due to regular exposure to the sun, an environmental gene. Some major characteristics, such as sex, are determined by the environment for some species. For instance, some turtles and other reptiles have temperature-dependent sex conclusion (TSD). TSD means that individuals develop into males if their eggs are incubated within a certain temperature range, or females at a dissimilar temperature range.
Geographic separation between populations tin lead to differences in the phenotypic variation between those populations. Such geographical variation is seen betwixt well-nigh populations and can be significant. One blazon of geographic variation, called a cline, can be seen as populations of a given species vary gradually across an ecological slope. Species of warm-blooded animals, for example, tend to accept larger bodies in the cooler climates closer to the world's poles, allowing them to amend conserve heat. This is considered a latitudinal cline. Alternatively, flowering plants tend to blossom at dissimilar times depending on where they are forth the slope of a mountain, known every bit an altitudinal cline.
If there is gene menstruation between the populations, the individuals volition probable prove gradual differences in phenotype forth the cline. Restricted gene flow, on the other hand, can lead to abrupt differences, even speciation.
Adaptive Evolution
Natural choice only acts on the population's heritable traits: selecting for benign alleles and thus increasing their frequency in the population, while selecting against deleterious alleles and thereby decreasing their frequency—a process known equally adaptive development. Natural choice does non act on private alleles, however, simply on unabridged organisms. An individual may comport a very beneficial genotype with a resulting phenotype that, for example, increases the ability to reproduce (fecundity), but if that same individual too carries an allele that results in a fatal childhood disease, that fecundity phenotype volition not be passed on to the next generation because the individual will non live to reach reproductive historic period. Natural selection acts at the level of the individual; it selects for individuals with greater contributions to the gene pool of the adjacent generation, known as an organism's evolutionary (Darwinian) fitness.
Fitness is often quantifiable and is measured by scientists in the field. However, it is not the absolute fitness of an individual that counts, just rather how it compares to the other organisms in the population. This concept, called relative fitness, allows researchers to make up one's mind which individuals are contributing boosted offspring to the adjacent generation, and thus, how the population might evolve.
There are several ways selection tin affect population variation: stabilizing selection, directional option, diversifying selection, frequency-dependent selection, and sexual selection. Equally natural option influences the allele frequencies in a population, individuals can either become more than or less genetically similar and the phenotypes displayed can go more similar or more disparate.
Stabilizing Selection
If natural selection favors an boilerplate phenotype, selecting against extreme variation, the population will undergo stabilizing option (Figure 7). In a population of mice that live in the woods, for instance, natural selection is likely to favor individuals that all-time blend in with the wood floor and are less probable to exist spotted past predators. Assuming the basis is a fairly consequent shade of brown, those mice whose fur is most closely matched to that color will be well-nigh probable to survive and reproduce, passing on their genes for their brown coat. Mice that carry alleles that make them a bit lighter or a bit darker volition stand up out confronting the ground and be more likely to fall victim to predation. Equally a result of this selection, the population's genetic variance will decrease.

Figure seven. In stabilizing selection, an average phenotype is favored.
Directional Selection
When the environment changes, populations will oft undergo directional selection (Effigy 8), which selects for phenotypes at one terminate of the spectrum of existing variation. A archetype example of this type of selection is the development of the peppered moth in eighteenth- and nineteenth-century England. Prior to the Industrial Revolution, the moths were predominately light in colour, which allowed them to blend in with the lite-colored trees and lichens in their environment. Only as soot began spewing from factories, the copse became darkened, and the light-colored moths became easier for predatory birds to spot. Over fourth dimension, the frequency of the melanic grade of the moth increased because they had a college survival charge per unit in habitats affected by air pollution because their darker coloration blended with the sooty trees. Similarly, the hypothetical mouse population may evolve to take on a different coloration if something were to cause the forest floor where they live to modify color. The effect of this blazon of selection is a shift in the population's genetic variance toward the new, fit phenotype.

Figure eight. In directional selection, a change in the surround shifts the spectrum of phenotypes observed.
In science, sometimes things are believed to be true, then new data comes to calorie-free that changes our agreement. The story of the peppered moth is an instance: the facts behind the pick toward darker moths have recently been called into question. Read this article to learn more.
Diversifying Selection
Sometimes two or more than distinct phenotypes can each have their advantages and be selected for by natural choice, while the intermediate phenotypes are, on average, less fit. Known every bit diversifying selection (Effigy 9), this is seen in many populations of animals that have multiple male forms. Large, ascendant blastoff males obtain mates by fauna strength, while small males can sneak in for furtive copulations with the females in an alpha male'south territory. In this case, both the blastoff males and the "sneaking" males will be selected for, but medium-sized males, which can't overtake the alpha males and are too large to sneak copulations, are selected against. Diversifying selection can as well occur when environmental changes favor individuals on either end of the phenotypic spectrum. Imagine a population of mice living at the beach where at that place is light-colored sand interspersed with patches of tall grass. In this scenario, light-colored mice that blend in with the sand would exist favored, as well as night-colored mice that tin hide in the grass. Medium-colored mice, on the other hand, would non alloy in with either the grass or the sand, and would thus exist more than likely to exist eaten by predators. The consequence of this type of pick is increased genetic variance as the population becomes more diverse.

Figure 9. In diversifying pick, two or more extreme phenotypes are selected for, while the average phenotype is selected against.
Practice Question
Different types of natural selection can impact the distribution of phenotypes within a population (refer back to Figures 7, 8, and 9). In recent years, factories have become cleaner, and less soot is released into the environs. What affect exercise you lot recollect this has had on the distribution of moth color in the population?
Show Respond
Moths have shifted to a lighter colour.
Frequency-dependent Option
Another type of option, called frequency-dependent option, favors phenotypes that are either common (positive frequency-dependent selection) or rare (negative frequency-dependent selection). An interesting example of this type of selection is seen in a unique grouping of lizards of the Pacific Northwest. Male person common side-blotched lizards come in iii throat-color patterns: orange, blue, and yellow.

Effigy 10. A yellow-throated side-blotched lizard is smaller than either the blue-throated or orangish-throated males and appears a bit like the females of the species, allowing information technology to sneak copulations. (credit: "tinyfroglet"/Flickr)
Each of these forms has a dissimilar reproductive strategy: orange males are the strongest and can fight other males for access to their females; blue males are medium-sized and course strong pair bonds with their mates; and yellow males (Figure 10) are the smallest, and look a bit like females, which allows them to sneak copulations. Like a game of stone-paper-pair of scissors, orangish beats blue, blue beats yellow, and yellow beats orange in the contest for females. That is, the large, strong orangish males tin fight off the blue males to mate with the blue's pair-bonded females, the blue males are successful at guarding their mates confronting yellow sneaker males, and the yellow males can sneak copulations from the potential mates of the large, polygynous orange males.
In this scenario, orange males will be favored by natural selection when the population is dominated by blueish males, blue males will thrive when the population is mostly yellow males, and yellow males volition exist selected for when orange males are the most populous. As a issue, populations of side-blotched lizards wheel in the distribution of these phenotypes—in one generation, orange might be predominant, and so yellowish males will begin to rising in frequency. Once yellow males make upward a bulk of the population, blueish males will be selected for. Finally, when blue males become mutual, orangish males will once again be favored.
Negative frequency-dependent selection serves to increase the population'southward genetic variance past selecting for rare phenotypes, whereas positive frequency-dependent pick usually decreases genetic variance by selecting for common phenotypes.
Sexual Choice
Males and females of certain species are often quite dissimilar from i another in ways beyond the reproductive organs. Males are often larger, for instance, and brandish many elaborate colors and adornments, like the peacock'due south tail, while females tend to exist smaller and duller in decoration. Such differences are known as sexual dimorphisms (Figure 11), which arise from the fact that in many populations, peculiarly animal populations, there is more variance in the reproductive success of the males than there is of the females. That is, some males—often the bigger, stronger, or more than decorated males—get the vast majority of the full matings, while others receive none. This can occur because the males are better at fighting off other males, or because females will choose to mate with the bigger or more decorated males. In either instance, this variation in reproductive success generates a strong selection pressure amid males to get those matings, resulting in the development of bigger body size and elaborate ornaments to get the females' attending. Females, on the other hand, tend to get a scattering of selected matings; therefore, they are more than probable to select more desirable males.
Sexual dimorphism varies widely among species, of grade, and some species are even sex-part reversed. In such cases, females tend to have a greater variance in their reproductive success than males and are correspondingly selected for the bigger trunk size and elaborate traits usually characteristic of males.

Effigy 11. Sexual dimorphism is observed in (a) peacocks and peahens, (b) Argiope appensa spiders (the female spider is the big ane), and in (c) wood ducks. (credit "spiders": modification of work past "Sanba38"/Wikimedia Eatables; credit "duck": modification of work past Kevin Cole)
The selection pressures on males and females to obtain matings is known every bit sexual selection; it tin can outcome in the development of secondary sexual characteristics that practise not benefit the individual's likelihood of survival but assist to maximize its reproductive success. Sexual selection can be and so potent that it selects for traits that are actually detrimental to the individual's survival. Think, once more, almost the peacock's tail. While information technology is cute and the male with the largest, most colorful tail is more likely to win the female, it is non the most practical appendage. In addition to beingness more visible to predators, it makes the males slower in their attempted escapes. There is some evidence that this risk, in fact, is why females like the big tails in the offset place. The speculation is that large tails carry take a chance, and only the best males survive that risk: the bigger the tail, the more than fit the male. This idea is known as the handicap principle.
The good genes hypothesis states that males develop these impressive ornaments to show off their efficient metabolism or their ability to fight disease. Females and then choose males with the most impressive traits considering it signals their genetic superiority, which they will and so pass on to their offspring. Though information technology might be argued that females should not exist picky considering it will likely reduce their number of offspring, if better males father more fit offspring, information technology may be beneficial. Fewer, healthier offspring may increase the chances of survival more than many, weaker offspring.
In 1915, biologist Ronald Fisher proposed some other model of sexual selection: the Fisherian runaway model, which suggests that selection of sure traits is a issue of sexual preference.
In both the handicap principle and the good genes hypothesis, the trait is said to be an honest point of the males' quality, thus giving females a way to detect the fittest mates—males that volition pass the all-time genes to their offspring.
No Perfect Organism
Natural selection is a driving force in evolution and tin generate populations that are better adapted to survive and successfully reproduce in their environments. But natural option cannot produce the perfect organism. Natural option can only select on existing variation in the population; it does non create anything from scratch. Thus, it is express by a population's existing genetic variance and whatever new alleles arise through mutation and gene flow.
Natural pick is also limited because it works at the level of individuals, not alleles, and some alleles are linked due to their physical proximity in the genome, making them more probable to exist passed on together (linkage disequilibrium). Any given private may carry some beneficial alleles and some unfavorable alleles. It is the net consequence of these alleles, or the organism's fitness, upon which natural selection can deed. As a result, good alleles tin can be lost if they are carried past individuals that also take several overwhelmingly bad alleles; likewise, bad alleles tin can be kept if they are carried by individuals that take plenty good alleles to result in an overall fitness benefit.
Furthermore, natural selection can exist constrained past the relationships between unlike polymorphisms. One morph may confer a college fitness than another, but may not increase in frequency due to the fact that going from the less beneficial to the more beneficial trait would require going through a less beneficial phenotype. Think back to the mice that alive at the embankment. Some are light-colored and alloy in with the sand, while others are dark and blend in with the patches of grass. The dark-colored mice may be, overall, more fit than the calorie-free-colored mice, and at first glance, i might look the lite-colored mice be selected for a darker coloration. But remember that the intermediate phenotype, a medium-colored coat, is very bad for the mice—they cannot blend in with either the sand or the grass and are more likely to be eaten past predators. As a result, the light-colored mice would not be selected for a nighttime coloration because those individuals that began moving in that direction (began being selected for a darker coat) would be less fit than those that stayed light.
Finally, it is important to understand that not all development is adaptive. While natural selection selects the fittest individuals and ofttimes results in a more than fit population overall, other forces of evolution, including genetic migrate and gene flow, often do the reverse: introducing deleterious alleles to the population'southward gene pool. Evolution has no purpose—it is non changing a population into a preconceived platonic. It is simply the sum of the various forces described in this chapter and how they influence the genetic and phenotypic variance of a population.
Practice Questions
Give an case of a trait that may take evolved equally a result of the handicap principle and explain your reasoning.
List the ways in which evolution tin affect population variation and describe how they influence allele frequencies.
Evidence Answer
There are several means evolution can impact population variation: stabilizing option, directional option, diversifying selection, frequency-dependent selection, and sexual choice. Equally these influence the allele frequencies in a population, individuals can either go more or less related, and the phenotypes displayed can become more similar or more disparate.
Check Your Understanding
Answer the question(s) below to run into how well you lot sympathise the topics covered in the previous section. This brusque quiz doesnon count toward your class in the course, and you tin retake information technology an unlimited number of times.
Apply this quiz to check your agreement and decide whether to (1) study the previous section further or (2) move on to the adjacent section.
Source: https://courses.lumenlearning.com/suny-wmopen-biology2/chapter/the-evolution-of-populations/
0 Response to "The Evolution of Populations Reading Guide Answers"
Post a Comment